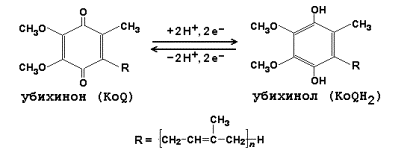1. Понятие о метаболизме (промежуточном обмене веществ). Основные функции метаболизма.
2. Автотрофные и гетеротрофные организмы. Роль фототрофов в преобразовании солнечной энергии.
3. Катаболические и анаболические пути обмена веществ. Взаимосвязь процессов катаболизма и анаболизма. Амфиболические пути.
4. Стадии катаболизма основных классов питательных веществ: локализация, конечные продукты, относительная энергоотдача. Понятие об общем и специфических путях катаболизма. Узловые (ключевые) метаболиты, примеры.
5. Окислительное декарбоксилирование пирувата: локализация в клетке, состав мультифермента, роль коферментов. Формулы коферментов: ТДФ, липоевая кислота, ФАД, НАД. Факторы, влияющие на активность пируватдегидрогеназного комплекса.
6. Цикл трикарбоновых кислот (ЦТК): локализация в клетке, последовательность реакций, ферменты, коферменты. Регуляторные ферменты ЦТК, их эффекторы. Биологическая роль ЦТК.
7. Амфиболическая функция ЦТК. Примеры реакций, пополняющих фонд оксалоацетата в митохондриях.
| Раздел 13.1 |
Общая характеристика обмена веществ. Две стороны (фазы) метаболизма.
|
13.1.1. Метаболизм (обмен веществ) – совокупность химических реакций, протекающих в живой клетке. Эти реакции протекают в определённой последовательности и тесно связаны между собой. Главные функции метаболизма в клетке:
13.1.2. Метаболизм включает множество метаболических путей. Метаболический путь - определенная последовательность ферментативных реакций в клетке. Промежуточные продукты реакций метаболического пути называются метаболитами. На каждой из стадий метаболического пути происходит небольшое химическое изменение метаболитов. В результате этих превращений исходная молекула превращается в конечный продукт метаболического пути.
13.1.3. Большинство метаболических путей являются линейными (рисунок 13.1, а), но встречаются и циклические метаболические пути (рисунок 13.1, б). Обычно метаболические пути имеют разветвления, в которых одни вещества выходят из цепи реакций данного пути, а другие, наоборот, включаются в него (рисунок 13.1, в).
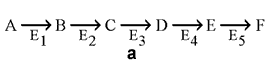 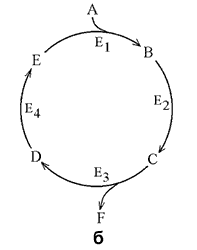 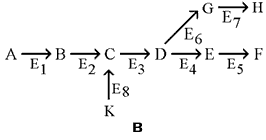
Рисунок 13.1. а. Линейный метаболический путь (схема). Предшественник А превращается в продукт F в результате пяти последовательных реакций. Продукт одной ферментативной реакции служит при этом субстратом следующей. б. Циклический путь (схема). Конечным продуктом такого пути является один из субстратов первой ферментативной реакции. Таким путём происходит окисление ацетильной группы до СО2 и Н2О в цикле Кребса. в. Разветвлённый метаболический путь (схема). Метаболит D выходит из цепи реакций данного пути, превращаясь в метаболит G, а вещество К, наоборот, включается в него.
Обратите внимание, что главные метаболические пути немногочисленны и одинаковы у большинства живых организмов. Именно эти пути представляют для нас наибольший интерес. Рассмотрению их будут посвящены разделы 13.3. и 13.4 настоящей темы.
13.1.4. Две стороны (фазы) метаболизма. Метаболизм складывается из двух противоположных сторон : катаболизма и анаболизма.
Катаболизм – это фаза, в которой происходит последовательное расщепление сложных молекул до более простых, таких, как СО2, вода и аммиак. Процессы катаболизма сопровождаются выделением энергии. Эта энергия частично аккумулируется в форме макроэргического соединения – аденозинтрифосфата (АТФ).
Анаболизм – это фаза метаболизма, в которой происходит образование (биосинтез) сложных молекул (белков, липидов, полисахаридов) из простых предшественников. Процессы биосинтеза протекают с затратой энергии. Источником этой энергии служит распад АТФ до АДФ и неорганического фосфата.
Метаболические пути, выполняющие как катаболическую, так и анаболическую функцию, называют амфиболическими.
13.1.5. Катаболическая и анаболическая фазы метаболизма тесно связаны между собой (рисунок 13.2):
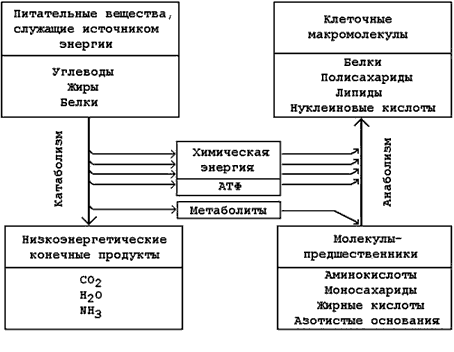 Рисунок 13.2. Взаимосвязь путей катаболизма и анаболизма | |
| Раздел 13.2 |
Стадии катаболизма питательных веществ.
|
13.2.1. Как уже было сказано (раздел 13.1), катаболизм – совокупность химических реакций превращения высокомолекулярных соединений в низкомолекулярные. Это сложный ферментативный процесс, в котором принято выделять три основные стадии (рисунок 13.3). Для каждой стадии запомните, какие вещества вступают и образуются, локализацию в организме и энергетическую значимость.
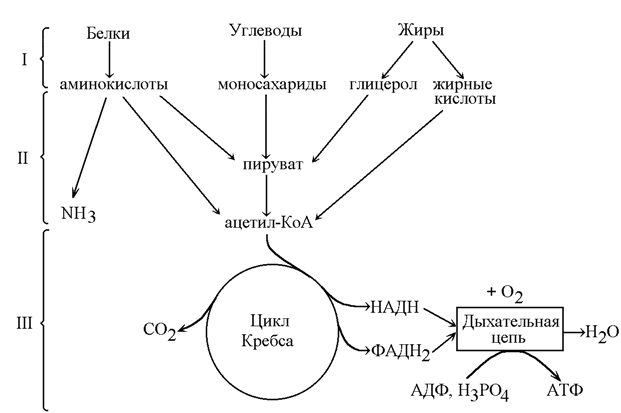
Рисунок 13.3. Три стадии катаболизма питательных веществ (обозначены римскими цифрами).
13.2.2. На первой стадии крупные биомолекулы расщепляются на составляющие их строительные блоки: полисахариды превращаются в пентозы и гексозы, жиры – в жирные кислоты, глицерол и другие компоненты, белки – в аминокислоты. Это происходит в желудочно-кишечном тракте, а также в лизосомах клетки. Реакции катализируют ферменты, относящиеся к классу гидролаз. Относительная энергоотдача составляет менее 1% всей высвобождаемой энергии.
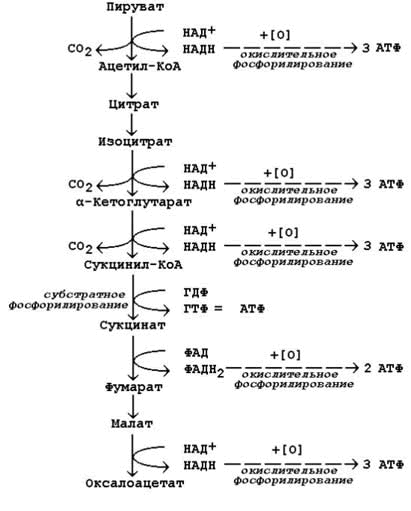
На второй стадии строительные блоки превращаются в более простые молекулы. Моносахариды, глицерол и большинство аминокислот расщепляются до одного и того же трёхуглеродного метаболита – пирувата. Это происходит в цитоплазме клеток. В дальнейшем пируват, а также жирные кислоты и некоторые аминокислоты окисляются до ацетильного остатка, связанного с коэнзимом А (ацетил-КоА). Эти реакции протекают уже в митохондриях клетки. Пируват и ацетил-КоА, находящиеся на пересечении нескольких метаболических путей, можно отнести кключевым или узловым метаболитам. Относительная энергоотдача второй стадии катаболизма около 20%; выделяемая энергия может быть частично аккумулирована в виде АТФ.
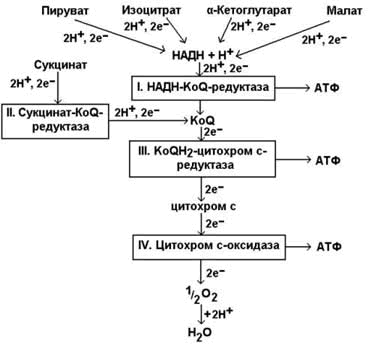
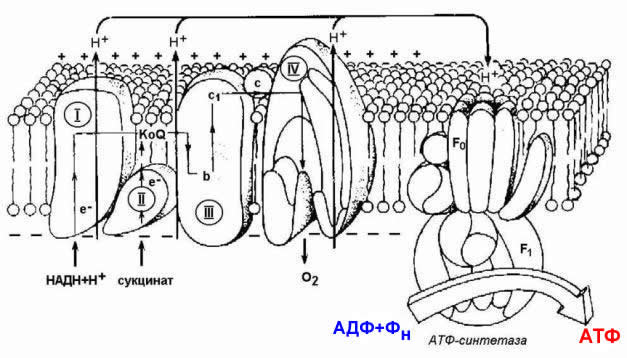
На третьей стадии происходит окисление ацетильной группы в цикле трикарбоновых кислот Кребса до СО2 и восстановленных форм коферментов НАД и ФАД. Эти коферменты окисляются в дыхательной цепи до Н2О; выделяемая энергия аккумулируется в АТФ. Все эти реакции протекают в митохондриях. Относительная энергоотдача третьей стадии - около 80%.
13.2.3. Различают общий и специфические пути катаболизма. К специфическим путям катаболизма веществ того или иного класса относят главным образом реакции первой и второй стадии катаболизма, которые для каждого класса могут существенно различаться. Третья стадия, одинаковая для всех классов питательных веществ, называется общим путём катаболизма.
| |
| Раздел 13.3 |
Окислительное декарбоксилирование пирувата.
|
13.3.1. Заключительной реакцией второй стадии катаболизма питательных веществ является окислительное декарбоксилирование пирувата.
Пируват подвергается окислительному декарбоксилированию до ацетил-коэнзима А (ацетил-КоА) в митохондриях клеток при участии пируватдегидрогеназного комплекса. Пируватдегидрогеназный комплекс – мультифермент, в состав которого входит 3 фермента:
1) пируватдекарбоксилаза (коферментом служит производное витамина В1 - тиаминдифосфат, ТДФ, формулу его см. в теме «Коферменты»);
2) трансацилаза (кофермент – липоевая кислота, ЛК, см. рисунок 13.4);
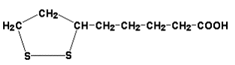
Рисунок 13.4. Липоевая кислота (витаминоподобное вещество).
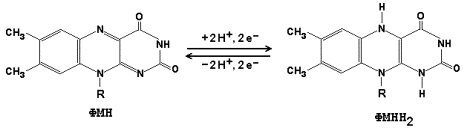
3) дигидролипоилдегидрогеназа (в качестве кофермента используется производное витамина В2 – флавинадениндинуклеотид, ФАД, формулу его см. в теме «Коферменты»).
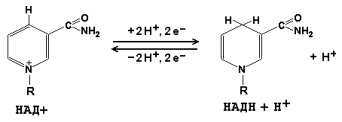
В реакции участвуют также два кофермента: коэнзим А (НSКоА, производное витамина В3) и никотинамидадениндинуклеотид (НАД, производное витамина РР), связанные при помощи нековалентных связей соответственно с трансацилазой и дигидролипоилдегидрогеназой.
13.3.2. Суммарное уравнение реакции, катализируемой пируватдегидрогеназным комплексом:
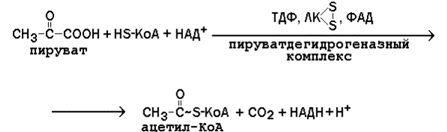
Схема работы этого мультиферментного комплекса представлена на рисунке 13.5.
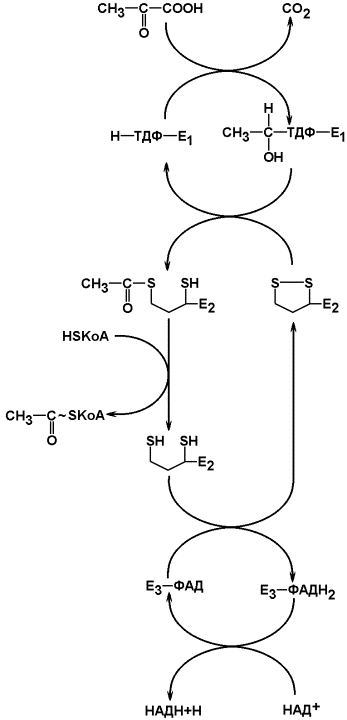
Рисунок 13.5. Окислительное декарбоксилирование пирувата (схема).
Ацетил-КоА затем подвергается окислению в цикле Кребса, а НАДН служит донором водорода для дыхательной цепи. Процесс является аэробным, т.к. конечный акцептор водорода НАДН – кислород. Скорость реакции снижается при накоплении в клетке ацетил-КоА, НАДН и АТФ, увеличивается – при возрастании концентрации АДФ. Регуляторным ферментом комплекса является пируватдекарбоксилаза.
Аналогично происходит окислительное декарбоксилирование α-кетоглутарата – одного из метаболитов цикла Кребса. Реакцию катализирует α-кетоглутаратдегидрогеназный комплекс, сходный с пируватдегидрогеназным комплексом (см. раздел 13.4, рис. 13.7, реакция 5).
| |
| Раздел 13.4 |
Цикл трикарбоновых кислот Кребса.
|
13.4.1. Реакции цикла Кребса относятся к третьей стадии катаболизма питательных веществ и происходят в митохондриях клетки. Эти реакции относятся к общему пути катаболизма и характерны для распада всех классов питательных веществ (белков, липидов и углеводов).
Главной функцией цикла является окисление ацетильного остатка с образованием четырёх молекул восстановленных коферментов (трёх молекул НАДН и одной молекулы ФАДН2), а также образование молекулы ГТФ путём субстратного фосфорилирования. Атомы углерода ацетильного остатка выделяются в виде двух молекул СО2.
13.4.2. Цикл Кребса включает 8 последовательных стадий, обращая особое внимание на реакции дегидрирования субстратов:
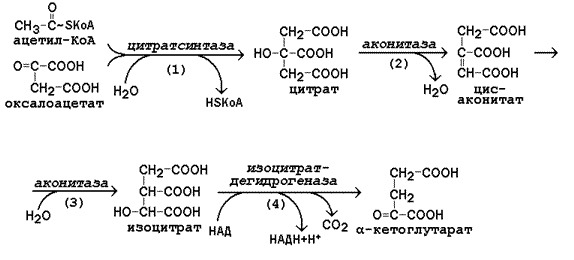
Рисунок 13.6. Реакции цикла Кребса, включая образование α-кетоглутарата
а) конденсация ацетил-КоА с оксалоацетатом, в результате которой образуется цитрат (рис.13.6, реакция 1); поэтому цикл Кребса называют также цитратным циклом. В этой реакции метильный углерод ацетильной группы взаимодействует с кетогруппой оксалоацетата; одновременно происходит расщепление тиоэфирной связи. В реакции освобождается КоА-SH, который может принять участие в окислительном декарбоксилировании следующей молекулы пирувата. Реакцию катализирует цитратсинтаза, это – регуляторный фермент, он ингибируется высокими концентрациями НАДН, сукцинил-КоА, цитрата.
б) превращение цитрата в изоцитрат через промежуточное образование цис-аконитата. Образующийся в первой реакции цикла цитрат содержит третичную гидроксильную группу и не способен окисляться в условиях клетки. Под действием фермента аконитазы идёт отщепление молекулы воды (дегидратация), а затем её присоединение (гидратация), но другим способом (рис.13.6, реакции 2-3). В результате данных превращений гидроксильная группа перемещается в положение, благоприятствующее её последующему окислению.
в) дегидрирование изоцитрата с последующим выделением молекулы СО2 (декарбоксилированием) и образованием α-кетоглутарата (рис. 13.6, реакция 4). Это – первая окислительно-восстановительная реакция в цикле Кребса, в результате которой образуется НАДН. Изоцитратдегидрогеназа, катализирующая реакцию, - регуляторный фермент, активируется АДФ. Избыток НАДН ингибирует фермент.
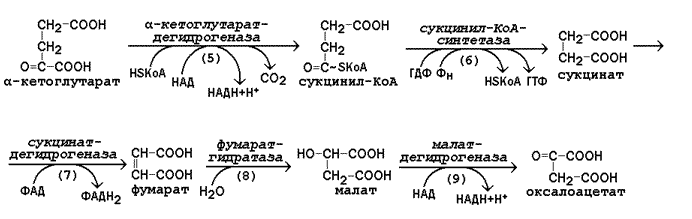
Рисунок 13.7. Реакции цикла Кребса, начиная с α-кетоглутарата.
г) окислительное декарбоксилирование α-кетоглутарата, катализируется мультиферментным комплексом (рис. 13.7, реакция 5), сопровождается выделением СО2 и образованием второй молекулы НАДН. Эта реакция аналогична пируватдегидрогеназной реакции. Ингибитором служит продукт реакции – сукцинил-КоА.
д) субстратное фосфорилирование на уровне сукцинил-КоА, в ходе которого энергия, освобождающаяся при гидролизе тиоэфирной связи, запасается в форме молекулы ГТФ. В отличие от окислительного фосфорилирования, этот процесс протекает без образования электрохимического потенциала митохондриальной мембраны (рис. 13.7, реакция 6).
е) дегидрирование сукцината с образованием фумарата и молекулы ФАДН2 (рис. 13.7, реакция 7). Фермент сукцинатдегидрогеназа прочно связан с внутренней мембраной митохондрии.
ж) гидратация фумарата, в результате чего в молекуле продукта реакции появляется легко окисляемая гидроксильная группа (рис. 13.7, реакция 8).
з) дегидрирование малата, приводящее к образованию оксалоацетата и третьей молекулы НАДН (рис.13.7, реакция 9). Образующийся в реакции оксалоацетат может вновь использоваться в реакции конденсации с очередной молекулой ацетил-КоА (рис. 13.6, реакция 1). Поэтому данный процесс носит циклический характер.
13.4.3. Таким образом, в результате описанных реакций подвергается полному окислению ацетильный остаток СН3-СО-. Количество молекул ацетил-КоА, превращаемых в митохондриях в единицу времени, зависит от концентрации оксалоацетата. Основные пути увеличения концентрации оксалоацетата в митохондриях (соответствующие реакции будут рассмотрены позднее):
13.4.4. Некоторые метаболиты цикла Кребса могут использоваться для синтеза структурных блоков для построения сложных молекул. Так, оксалоацетат может превращаться в аминокислоту аспартат, а α–кетоглутарат – в аминокислоту глутамат. Сукцинил-КоА принимает участие в синтезе гема – простетической группы гемоглобина. Таким образом, реакции цикла Кребса могут участвовать как в процессах катаболизма, так и анаболизма, то есть цикл Кребса выполняет амфиболическую функцию (см. 13.1).
| |
| Раздел 13.5 |
Примеры обучающих задач и методов их решения.
|
Вопросы для повторения.
1. Дайте определение понятия «метаболизм» и перечислите его основные функции.
2. Что понимают под процессами катаболизма и анаболизма в организме? Какая взаимосвязь существует между этими процессами?
3. Охарактеризуйте реакции первой стадии катаболизма питательных веществ в организме: укажите их локализацию, исходные вещества и образующиеся продукты, относительную энергоотдачу.
4. Охарактеризуйте реакции второй стадии катаболизма питательных веществ в организме: укажите их локализацию, исходные вещества и образующиеся продукты, относительную энергоотдачу.
5. Что понимают под специфическим и общим путями катаболизма питательных веществ? Какие стадии катаболизма к ним относят?
6. Напишите суммарную реакцию окислительного декарбоксилирования пирувата. Укажите ферменты, входящие в состав мультиэнзимного комплекса.
7. Напишите формулу тиаминдифосфата, укажите его роль в пируватдегидрогеназной реакции и витамин, входящий в его состав.
8. Напишите формулу липоевой кислоты, укажите её роль в пируватдегидрогеназной реакции. Как осуществляется связь липоевой кислоты с апоферментом?
9. Как и почему изменится активность пируватдегидрогеназного комплекса при увеличении отношения АТФ/АДФ в клетке? Назовите механизм регуляции.
10. Как и почему изменится скорость окислительного декарбоксилирования пирувата при увеличении отношения НАДН/НАД+ в клетке? Назовите механизм регуляции.
11. Напишите начальную реакцию цикла трикарбоновых кислот, назовите фермент. В чём заключается биологическая роль цикла Кребса?
12. Напишите реакцию окисления изолимонной кислоты в цикле Кребса, укажите фермент, кофермент и витамин, входящий в его состав.
13. Напишите реакцию окислительного декарбоксилирования α-кетоглутаровой кислоты в цикле Кребса, укажите фермент, коферменты и витамины, входящие в их состав.
14. Напишите реакцию субстратного фосфорилирования в цикле Кребса, назовите фермент. В чём заключается биологическая роль субстратного фосфорилирования?
15. Напишите реакцию окисления янтарной кислоты в цикле Кребса, укажите фермент, кофермент и витамин, входящий в его состав.
16. Напишите реакцию окисления яблочной кислоты в цикле Кребса, укажите фермент. Почему цикл Кребса называют общим путём катаболизма?
17. Перечислите субстраты, подвергающиеся дегидрированию в реакциях цикла Кребса. Сколько и какие восстановленные формы коферментов образуются в этих реакциях?
18. Что понимают под амфиболической функцией цикла Кребса? Напишите анаплеротическую реакцию цикла Кребса.
19. Как и почему изменится скорость реакций ЦТК при уменьшении отношения АТФ/АДФ в клетке? Назовите механизм регуляции.
20. Как и почему изменится скорость реакций ЦТК при уменьшении отношения НАДН/НАД+ в клетке? Назовите механизм регуляции.
| |